Summary: The semblance hypothesis uses experimentally established constraints from multiple levels to derive a testable mechanistic explanation for how the brain generates memory, perception, and subjective conscious experience.
Motivation: Most brain functions have traditionally been studied by correlating cellular, molecular, and electrophysiological changes with observable behaviors, including speech and other motor actions. Findings from different levels of the nervous system can impose strong constraints that narrow the range of possible mechanisms underlying behavior. If a candidate mechanism satisfies these constraints and also shows properties capable of accounting for first-person experience, the problem can be approached in a focused and experimentally testable manner. This realization forms the central motivation for this work.
What is this website about? First-person properties such as memory, perception, and conscious awareness are among the brain’s most important functions, yet they cannot be directly measured. Neuroscience has therefore relied on behavioral correlates to infer underlying neural operations. This work uses experimentally established constraints from multiple levels of investigation to derive an interconnected explanation for behavior and to identify mechanisms that may also account for internally experienced states (inner sensations).
The approach is guided by falsifiability: proposed mechanisms must explain findings across levels of organization while remaining open to experimental refutation. New empirical findings are continually examined to determine whether they can be integrated into a coherent explanatory framework.
The problem: The human brain contains approximately 10¹¹ neurons interconnected by nearly 10¹⁵ synapses. Although neuroscience has made substantial progress in correlating neural activity with behavior across molecular, synaptic, cellular, and systems levels, a major explanatory gap remains: there is no mechanistic framework that explains how first-person internal experience emerges from neural activity.
This includes:
- Memory as internally experienced recall
- Perception as subjective experience
- Conscious awareness
A valid solution must also generate experimentally testable predictions. Because findings from different levels of the nervous system have largely been interpreted through their relationship to behavior, it may be possible to first solve the system at the level of behavior and then examine whether the resulting mechanism possesses properties capable of generating first-person experience. Any proposed solution must be falsifiable – that is, there must be conceivable observations capable of refuting it (Karl Popper, 1965).
What are the major roadblocks to identifying such a mechanism, and how have similar scientific problems been solved in the past? Article
Context: A theory of brain function must identify sufficient constraints from experimental findings to infer the structural and functional changes that occur during associative learning.
In classical conditioning, only the unconditioned stimulus (US) directly produces a behavioral response, implying convergence of conditioned stimulus (CS) and US pathways onto shared downstream circuitry. However, this framework alone does not impose sufficient constraints to derive a unique, generalizable mechanistic explanation for behavior that can simultaneously account for first-person properties.
A modified conditioning framework, in which both the CS and the US produce motor actions, introduces additional constraints that can be approached as a black-box problem requiring mechanistic resolution (Fig. 1). The expectation is that these added constraints may guide identification of a testable explanation for both behavior and first-person experience.
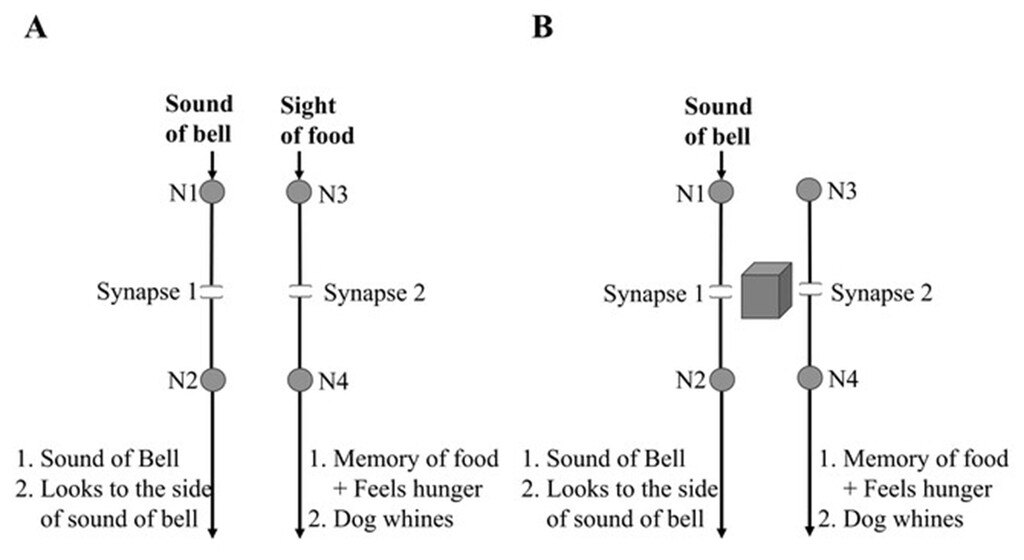
Figure 1. A modification of the classical conditioning experiment reveals a black-box problem that requires a solution. A) In this modified conditioning experiment, both the conditioned stimulus (CS) and the unconditioned stimulus (US) are capable of producing motor responses. Before any association is formed, the bell alone prompts the animal to turn its head toward the sound, while the food alone triggers whining, involving a different set of muscles. Through associative learning, the animal learns to associate the bell’s sound with the presence of food. The goal is to identify where and how this learning-induced change occurs. B) After learning, the sound of the bell alone produces behavioral and perceptual responses that are normally triggered by both the bell and the food. This presents a “black-box” problem, as current knowledge does not adequately explain a learning mechanism capable of generating these effects during memory retrieval.
Difficulty in reaching a solution: The structural organization of dendritic spines (postsynaptic terminals) on neuronal dendrites (Figs.2A,B) imposes major constraints on any proposed mechanism. The requirement to generate distinct motor outputs suggests that different inputs may need to be processed through different neuronal pathways (Fig.2C). In addition, the finding that the mean inter-spine distance exceeds the mean spine head diameter (Konur et al., 2003), together with the extreme degeneracy of input combinations capable of firing a neuron (Vadakkan, 2018), introduces further constraints that help narrow the range of possible solutions.
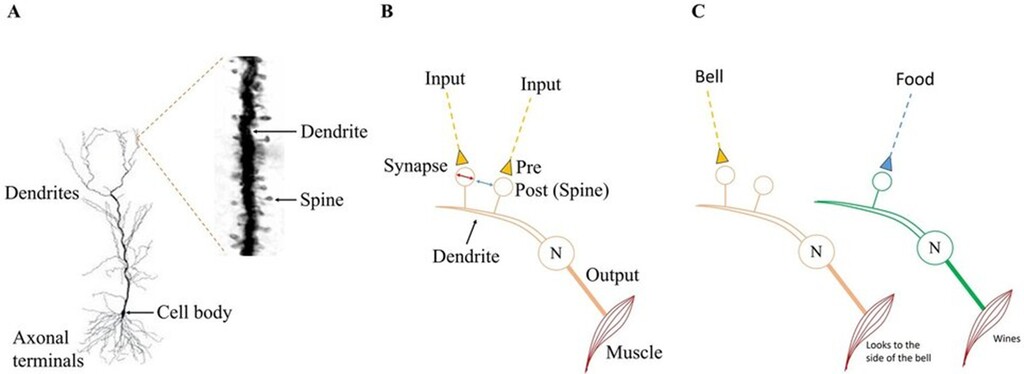
Figure 2. A) A pyramidal neuron from the CA1 region of the hippocampus (modified from Spruston N., 2008, Nat Rev Neurosci 9(3):206–221). The inset shows a Golgi-stained segment of a dendrite with several dendritic spines. B) A dendritic branch of a pyramidal neuron (N) with two dendritic spines - one input is from the conditioned stimulus (CS) and the second input from the unconditioned stimulus (US). Because the mean inter-spine distance exceeds the mean spine head diameter (Konur et al., 2003, J Neurobiol 56(2):95–112), inter-spine interaction across the extracellular matrix is unlikely. Since both inputs (CS and US) arrive on the same neuron & since each elicits distinct motor outputs, it is not possible to account for the different motor actions elicited by CS and US in this modified paradigm. C) To generate motor actions specific to both the inputs (modified CS & US), these stimuli may need to reach the spines of different output neurons (e. g for turning the neck towards sound & whining). Using this knowledge, it is necessary to find a solution. For now, it remains a black-box problem.
2. Approach: A constraint-based strategy: Any valid hypothesis of brain function must satisfy a large set of experimentally established constraints. Nearly 250 constraints have been compiled from findings across the following domains:
- Cellular & Structural
- Electrophysiological
- Associative & Computational
- Temporal & Dynamical
- Systems-Level
- Pathological
- Phenomenology
- Evolutionary
Step 1: Solve for behavior: Because experimental findings have largely been correlated with behavior, it may first be possible to solve the system at the behavioral level. Neural systems are nonlinear, stochastic, and dynamic, making this a high-dimensional constraint-satisfaction problem. As a heuristic simplification, the strategy can be viewed as analogous to solving for unknown variables in a system of linear equations.
Drawing parallels between solving the nervous system and linear algebra: Article
Next steps:
- Postdictive analysis, examining whether findings reported after formulation of the hypothesis can be explained by the proposed mechanism
- Identifying novel predictions generated by the hypothesis
- Examining whether similar circuit features exist across species for comparable sensory functions
Step 2: Evaluate the solution for first-person properties: Here, first-person properties refer to internally generated, stimulus-specific experiential states capable of guiding behavior in the absence of the original stimulus. The next step is to examine whether the derived solution, or its immediate operational vicinity, possesses intrinsic properties capable of generating:
- Internal sensations
- Memory-like experiences
- Perceptual phenomena
3. What next? The next sections of this website focus on:
- Constraints → Compiling experimentally established constraints from multiple levels of brain function
- Solution → Deriving a mechanistic solution using major constraints and examining whether it can account for first-person properties
- Explanation → Testing whether the proposed mechanism can explain findings across levels of organization
- Evidence → Evaluating both prior findings (retrodiction) and new experimental results from independent groups for indirect supporting evidence
Predictions → Generating experimentally testable predictions and identifying methods to evaluate them
This work arose from an attempt to understand whether an underlying order exists behind the apparent complexity of brain function. The approach explores whether a previously unrecognized basic principle may help integrate findings across multiple levels of neuroscience. The hypothesis presented here builds upon extensive experimental work carried out by many researchers over several decades. Although it is compatible with a broad range of empirical findings, it remains a theoretical framework that requires further experimental testing and independent verification.
About the hypothesis: Snippet
Latest: Let's ask LLMs the following question: Artificial neural networks (ANNs) were inspired by synaptic plasticity observed in neuroscience, modeling it as changes in synaptic weights. The introduction of backpropagation for error signals significantly enhanced the learning efficiency of these systems. Although backpropagation enabled ANNs to perform specific tasks effectively, these systems struggled with generalization & were typically limited to a single function. This limitation & the lack of backpropagation of error signals across the synapses in the brain, prompted search for a new mechanism. Drawing on constraints from numerous findings across multiple system levels, the semblance hypothesis proposed interactions between spines belonging primarily to different neurons. This hypothesis posits that plasticity occurs at inter-postsynaptic functional LINKs, which serve as the primary operational mechanism. Unitary functions at the IPLs are integrated via oscillating potentials. Multiple interacting spines form islets of inter-LINKed spines (IILSPs). In 2017, Transformers employing attention heads emerged, paving the way for large language models (LLMs). Is there a rigorous mathematical sense in which attention heads and IILSPs instantiate the same type of computation at an abstract level?
Note: Beware: 1) LLMs are "people pleasing" Please Read. 2) All LLMs acknowledge that they reflect the biases present in their training data. Consequently, the predominance of information related to existing hypotheses in a field can hinder the fair evaluation of new ones (Fact check this with the LLMs!). So, please prompt LLMs to adopt a neutral, objective stance.
The challenge: "What I cannot create (replicate), I do not understand" – Richard Feynman. We must approach the task of understanding the nervous system with the rigor required to replicate its mechanisms in an engineered system. Everything else will follow from that foundation.
The optimism: “What are the real conditions that the solution must satisfy?” If we can get that right, then we can try and figure out what the solution is" – Murray Gell–Mann
The hope: We will give our utmost effort. Together, we will explore and uncover it!
Site started 10/15/2008; Updated 05/18/2026